Мастер-класс с пошаговыми фото
Если вы задумались над вопросом о том, чем бы новым и интересным занять вашего маленького мастера, то предлагаем вам...
Различные виды рецепторов кожи (схема) 1 -- свободные нервные окончания из роговицы глаза; 2 -- осязательные пластинки Меркеля; 3 -- осязательные тельца Мейсснера; 4 -- нервное сплетение волосяной луковицы; 5 -- концевая колба Краузе; 6 -- тельце Гольджи--Маццони
Болевые рецепторы (ноцицепторы) - свободные нервные окончания. Кожные нервные сплетения состоят из двух слоев и из верхнего слоя к клеткам эпидермиса отходят в виде четок тонкие конечные волоконца. Ветви одного нервного волокна образуют в коже сеть площадью в 1 см2. Сети, возникшие при ветвлении разных волокон, настолько тесно друг с другом переплетаются, что сигналы осязания и боли идут сразу по нескольким нервным путям. Подобные сплетения встречаются всюду -- в коже, слизистых оболочках, во внутренних органах. Наибольшее количество ноцицепторов удается обнаружить в коже и роговице. В подмышечной и паховых областях, а также в надпочечных ямках число болевых точек равно 200 на 1 см2. На коже и на слизистых оболочках можно найти участки, не воспринимающие боли при уколе, щипке, сильном давлении.

Болевые рецепторы и нервные волокна кожи человека (схема)
В последние годы удалось обнаружить тонкие волоконца, связывающие свободные нервные окончания с рецепторами прикосновения, тепла и холода. Они получили название волокон Тимофеева. Наличием этих волокон можно объяснить тот факт, что усиление давления может вызвать чувство боли. Для того чтобы вызвать чувство прикосновения в осязательной точке, надо приложить давление 2--3 г на 1 мм2. А для того чтобы в той же точке вызвать боль, необходимо давление в 200 г на 1 мм2.
Тельца Фатер-Пачини (пластинчатые тельца) -- капсулированные рецепторы давления (барорецепторы) в округлой многослойной капсуле. Располагаются в дерме, чаще - на границе дермы и гиподермы. Являются быстроадаптирующимися (реагируют только в момент начала воздействия), то есть регистрируют силу давления. Обладают большими рецептивными полями, то есть представляют грубую чувствительность. Распределены в коже области пальцев, наружных половых органов (кроме того присутствуют в стенке мочевого пузыря, капсуле внутренних органов и др.) Пластинчатые тельца - самые крупные из всех инкапсулированных нервных окончаний. Они овальные, достигают 3-4 мм в длину и 2 мм в толщину. Для них характерно наличие многослойной пластинчатой соединительнотканной оболокой (наружной колбы), богатой гемокапиллярами. Под соединительнотканной оболочкой лежит наружная луковица, состоящая из 1060 концентрических пластинок, образованных уплощенными гексагональными периневральными эпителиоидными клетками. Войдя в тельце, нервное волокно теряет миелиновую оболочку. Внутри тельца оно окружено лимфоцитами, которые формируют внутреннюю луковицу.
Тельца Мейсснера -- рецепторы давления (барорецепторы), расположенные в дерме. Представляют собой слоистую структуру с нервным окончанием, проходящим между слоями. Являются быстроадаптирующимися. Обладают малыми рецептивными полями, то есть представляют тонкую чувствительность. Расположены в сосочковом слое кожи пальцев, губ, век, половых органов. Они имеют диаметр около 100 мкм, снаружи окружены соединительнотканной капсулой. В составе этих телец нейроглиальные клетки образуют внутреннюю колбу вокруг концевого утолщения чувствительного нервного волокна, которое располагается параллельно поверхности кожи.
Тельца Гольджи-Маццони (луковицеобразные тельца) - рецепторы давления (барорецепторы) - инкапсулированные чувствительные нервные окончания, состоящие из разветвления чувствительного нервного волокна, глиальной внутренней колбы и соединительнотканной капсулы; встречается в коже, соединительной оболочке глазного яблока, брюшине, клиторе, головке полового члена, коже губ и краев рта, а также в других покровах тела.
Тельца (клетки) Меркеля -- некапсулированные рецепторы давления (барорецепторы). Являются медленноадаптирующимися (реагируют на всей продолжительности воздействия), то есть регистрируют продолжительность давления. Обладают малыми рецептивными полями. Они принимают участие в восприятии прикосновений, поскольку тесно связаны с сетевидными концевыми разветвлениями чувствительных нервов. Кроме того, клетки Меркеля синтезируют специфические для нервных клеток маркеры (нейрофиламенты, нейрональные клеточные адгезионные молекулы и др.). На основании присутствия в цитоплазме клеток нейропептидов они относятся к диффузной эндокринной системе. Мет-энкефалин, вырабатываемый клетками Меркеля, стимулирует иммунные реакции организма.
Рецепторы волосяных луковиц -- реагируют на отклонение волоса.
Окончания Руффини -- рецепторы растяжения. Являются медленноадаптирующимися, обладают большими рецептивными полями. Реагируют на смещение кожи, тепло (терморецепторы) и на давление. Лежат в глубоких слоях кожи, например, подошвы стопы. Диаметр -- до 1 мм. Афферентное волокно образует наподобие кустика из немиелинизированных веточек, которые оканчиваются колбообразными терминалями (вздутиями, окруженными леммоцитами). Окончания плотно прилегают к фибробластам и коллагеновым волокнам, формирующим основу тельца. Соединительнотканная капсула хорошо выражена.
Концевые колбы Краузе - рецепторы, реагирующий на холод (терморецепторы). Обнаруживаются в конъюнктиве, языке, наружных половых органах. Диаметр -- до 150 мкм. Сферические по форме, имеют тонкую капсулу, многочисленные разветвления афферентного окончания располагаются в виде колбы.
Общее число температурных точек на поверхности кожи взрослого человека равно приблизительно 280 тыс., причем 30 тыс. приходится на долю точек, воспринимающих тепло. На поверхности тела температурные точки распределены весьма неравномерно. Наиболее чувствительны к температурным раздражениям веки глаза, грудные железы, спина. Область лба мало чувствительна к теплу и очень восприимчива к холоду. Мало чувствительны к резким тепловым раздражениям кожа головы, нижних конечностей, слизистая оболочка полости рта и языка.
Кожа обильно снабжена нервами и представляет собой большое рецепторное поле, воспринимающее раздражения, которые поступают из внешней и внутренней среды.
Ввиду наличия богатого и разнообразного нервного аппарата кожа играет важную роль в жизнедеятельности организма.
Нервный аппарат кожи состоит из нервных волокон и нервных окончаний, свободных или инкапсулированных. Они расположены преимущественно в дерме и эпидермисе; значительно меньше их в гиподерме.
Нервные стволы, проникающие в кожу, образуют нервное сплетение в гиподерме. От этого сплетения отходят нервы в дерму, образуя в ней новые сплетения. От нервных сплетений гиподермы и дермы отходят нервные веточки к волосяным мешочкам, сальным и потовым железам, мышцам.
В шиловидном слое эпидермиса находятся специальные нервные аппараты - клетки Меркеля, воспринимающие тактильную чувствительность, и свободные окончания осевых цилиндров в виде заострений и пуговчатых утолщений, воспринимающих болевую чувствительность.

В сосочковом слое дермы расположены нервные окончания, так называемые колбы Краузе и тельца Мейсснера. Колбы Краузе воспринимают чувство холода, их много в коже кистей, слизистых оболочках, в области клитора, головки полового члена и внутреннего листка крайней плоти.

Тельца Мейсснера, воспринимающие прикосновение, в большом количестве находятся в коже ладонной и боковой поверхностей пальцев.

В коже насчитывается около 500 000 клеток Меркеля и телец Мейсснера.
В подкожной жировой клетчатке расположены нервные приборы, воспринимающие чувство тепла - тельца Руффини и ощущение глубокого давления - тельца Фатера - Пачини.


На 1 см 2 кожи приходится до 200 болевых рецепторов, 20 тактильных 12 Холодовых и 2 тепловых.
Слизистая оболочка полости рта выстлана многослойным плоским эпителием, который значительно отличается от эпидермиса кожи и почти не имеет рогового, блестящего и зернистого слоев. Эпителий фактически состоит из базального и шиловидного слоев.
Шиловидный слой в верхней части состоит из нескольких плоских, как бы сдавленных клеток. Ороговения эпителия в обычных условиях не происходит, за исключением участков на передней поверхности твердого неба и верхушек нитевидных сосочков языка. Из-за отсутствия трех слоев эпителия мелкие многочисленные кровеносные сосуды слизистой оболочки полости рта легко просвечиваются и придают ей красный цвет.
Строма (дерма) здесь имеет такое же строение, как и в коже, но в ней больше слюнных, слизистых, сальных, серозных (белковых) и смешанных желез.
«Кожные и венерические болезни»,
А.А.Студницин, Б.Г.Стоянов
Рецепторные (чувствительные) нервные окончания воспринимают сигналы из внешней среды (экстерорецепторы) и внутренних органов (интерорецепторы). В зависимости от природы раздражения, регистрируемого рецепторами, они подразделяются в соответствии с физиологической классификацией, на механорецепторы, хеморецепторы, терморецепторы и болевые рецепторы (ноцицепторы). В специализированных органах чувств (орган вкуса, обоняния, зрения, равновесия и слуха) имеются особые рецепторные клетки, которые воспринимают соответствующие раздражения.
Морфологическая классификация чувствительных нервных окончаний основана на особенностях их структурной организации. В соответствии с этой классификацией различают свободные и несвободные чувствительные нервные окончания; последние включают инкапсулированные и неинкапсулированные окончания (рис. 8.18).
Рис. 8.18. Рецепторные (чувствительные) нервные окончания (по Rohen J.W., Lutjen-Drecoll E. 1982, с изменениями). 1 - свободные нервные окончания (СНО) образованы терминальными ветвлениями дендрита чувствительного нейрона, идущего в составе нервного волокна (НВ). Несвободные инкапсулированные нервные окончания (2-4) образованы ветвлениями дендрита, окруженными леммоцитами, в совокупности с которыми они образуют структуру, называемую внутренней колбой (ВК). Снаружи окончания покрыты соединительнотканной капсулой (СТК). 2 - колба Краузе, 3 - осязательное тельце (Мейснера), 4 - пластинчатое тельце (Фатер-Пачини).
Свободные чувствительные нервные окончания состоят только из терминальных ветвлений дендрита чувствительного нейрона, Они встречаются в эпителии, а также в соединительной ткани. Проникая в эпителиальный пласт, нервные волокна утрачивают миелиновую оболочку и нейролемму, а базальная мембрана их леммоцитов сливается с эпителиальной. Свободные нервные окончания обеспечивают восприятие температурных (тепловых и холодовых), механических и болевых сигналов (рис. 8.19).

Рис. 8.19. Свободные нервные окончания в соединительной ткани, представленные концевыми ветвлениями дендритов чувствительных нейронов.
Несвободные чувствительные нервные окончаний содержат все компоненты нервного волокна. Они разделяются на инкапсулированные (имеющие особую соединительнотканную капсулу) и неинкапсулированные.
Несвободные неинкапсулированные нервные окончания состоят из ветвлений дендритов, окруженных леммоцитами. Они встречаются в соединительной ткани кожи (дерме), а также собственной пластинки слизистых оболочек.
Несвободные инкапсулированные нервные окончания весьма разнообразны, но имеют единый общий план строения: их основу составляют ветвления дендрита, которые непосредственно окружены леммоцитами и снаружи покрыты особой соединительнотканной капсулой (см. рис. 8.18.). К этому виду нервных окончаний относят пластинчатые тельца (Фатер-Пачини), осязательные тельца (Мейснера) тельца Руффини, колбы Краузе, нервно-мышечные веретена и нервно-сухожильные веретена (сухожильные органы Гольджи).
Пластинчатые тельца (Фатер-Пачини) встречаются в соединительной ткани внутренних органов и кожи (рис. 8.20.). Они имеют вид округлых образований диаметром 1-5мм, воспринимают давление и вибрацию. Структурными компонентами тельца являются:
1) внутренняя колба (луковица), образованная видоизмененными уплощенными леммоцитами, в которую проникают одно или несколько нервных волокон, имеющих прямой ход;
2) наружная колба - слоистая соединительнотканная капсула, состоящая из фибробластов и коллагеновых волокон, образующих 10-60 концентрических пластин, между которыми имеется жидкость.
При деформации пластин капсулы давление передается на нервное окончание, что вызывает деполяризацию его мембраны.
 А А
|  Б Б
|
Рис. 8.20. На электронной микрофотографии изображены: Тельца Фатер-Пачини (РС) – инкапсулированный рецептор. Капсула состоит из параллельно расположенной пластинки (по-видимому, производной Шванновской клетки), коллагеновых волокон. В центре капсулы раположены ветвления одного немиелинизированного нервного волокна (х100).
Осязательные тельца (Мейснера) расположены преимущественно в сосочковом слое дермы, имеют эллипсоидную форму и небольшие размеры (около 50-120 мкм) (рис. 8.21.). Их внутренняя колба состоит из плоских глиальных клеток, лежащих перпендикулярно длинной оси тельца, между которыми располагаются веточки дендритов. Между глиальными клетками проникают коллагеновые фибриллы, связанные с базальным слоем эпителия. Капсула тонкая, переходит в периневрий.
 А А
|  Б Б
|
Рис. 8.21. Тельца Мейснера. Инкапсулированный рецептор (М), находящийся в коже губ, гениталий и т.д. Имеет овальную форму, располагается в коже прямо под эпидермисом (Е). Рецептор состоит из капсулы, образованной коллагеновыми волокнами, Шванновскими клетками (х320). (Б метод импрегнации х150). В капсуле находятся несколько безмякотных нервных окончаний, которые обильно ветвятся.).
Тельца Руффини лежат в соединительнотканной части кожи и капсулах суставов; они воспринимают давление и имеют вид веретеновидных структур длиной до 1-2 мм. Внутреннюю колбу образуют глиальные клетки, между которыми располагаются многочисленные ветвящиеся терминали дендритов с расширениями на концах. Капсула хорошо выражена, образована коллагеновыми волокнами.
Колбы Краузе - мелкие (40-150 мкм) округлые тельца, являющиеся механорецепторами и, возможно, холодовыми рецепторами. Они расположены в соединительной ткани сосочкового слоя дермы и собственной пластинке слизистой оболочки полости рта, надгортанника, в конъюнктиве глаза. Внутренняя колба образована уплощенными глиальными клетками, между которыми тонкие веточки дендрита образуют сплетение в виде клубочка. Капсула состоит из плоских клеток, являющихся продолжением периневрия.
Нервно-мышечные веретена - рецепторы растяжения волокон поперечнополосатых мышц - сложные инкапсулированные нервные окончания, обладающие как чувствительной, так и двигательной иннервацией. Число веретен в мышце зависит от ее функции и тем выше, чем более точными движениями она обладает. Нервно-мышечное веретено (рис. 8.22, 8.23.) имеет длину 0,5-7 мм и располагается параллельно ходу волокон мышцы, называемых экстрафузальными (от лат. extra - вне и fuso - веретено, т.е. расположенными за пределами веретена). Веретено покрыто тонкой соединительнотканной капсулой (продолжением периневрия), внутри которой находятся тонкие поперечнополосатые интрафузальные мышечные волокна двух видов:
Волокна с ядерной сумкой - в расширенной центральной части которых содержатся скопления ядер (1-4 волокна/веретено);
Волокна с ядерной цепочкой - более тонкие с расположением ядер в виде цепочки в центральной части (до 10 волокон/веретено).
Чувствительные нервные волокна образуют кольцеспиральные окончания на центральной части ишрафузальных волокон обоих типов и гроздьевидные окончания у краев волокон с ядерной цепочкой.
Двигательные нервные волокна - тонкие, образуют мелкие нервно-мышечные синапсы по краям интрафузальных волокон, обеспечивая их тонус.

Рис. 8.22. Нервно-мышечное веретено. 1 - общий вид веретена, располагающегося между экстрафузальными мышечными волокнами (ЭФМЗ) и образованного интрафузальными мышечными волокнами (ИФМВ), которые окружены соединительнотканной капсулой (СТК). 2 - детали строения веретена, содержащего два вида ИФМВ: волокна с ядерной сумкой (ВЯС) и волокна с ядерной цепочкой (ВЯЦ). Чувствительные нервные волокна образуют кольцеспиральные окончания (КСО) на центральной части ИФМВ обоих типов и гроздьевидные окончания (ГВО) у краев ВЯЦ. Нервно-мышечное веретено содержит также двигательные нервные волокна и образованные ими нервно-мышечные синапсы по краям ИФМВ (не показаны).
 А А
|  Б Б
|
Рис. 8.23. Нервно-мышечное веретено N-нервное волокно, С – капсула.А – продольный срез (х320), Б – поперечный.
Нервно-сухожильные веретена (сухожильные органы Гольджи) - рецепторы растяжения - веретеновидные инкапсулированные структуры длиной около 0.5-1мм, располагающиеся в области соединения волокон поперечнополосатых мышц с коллагеновыми волокнами сухожилий. Каждое веретено образовано капсулой из плоских фиброцитов (продолжение периневрия), которая охватывает группу сухожильных пучков, оплетенных многочисленными терминальными веточками нервных волокон, частично покрытых леммоцитами. Возбуждение рецепторов возникает при растяжении сухожилия во время мышечного сокращения.
СПИСОК ЛИТЕРАТУРЫ
ОСНОВНАЯ ЛИТЕРАТУРА
1. Заварзин А.А. Основы сравнительной гистологии. Л. ЛГУ, 1985.- 397 с.
2. Гистология. Под ред. Ю.И. Афанасьева, Н.А.Юриной.- М.: Медицина, 1989.- 670 с.
3. Хэм А., Кормак Д. Гистология. (в 5 томах). М.: Мир. 1982.
4. Антипчук Ю.П. Гистология с основами эмбриологии. - М:Просвещение. 1983.- 240 с.
5. Антипчук Ю.П. Гiстологiя з основами ембрiологii. – K.: Вища школа. 1976.- 141 с.
6. Волкова О.В., Елецкий О.Н. Основы гистологии с гистологической техникой. –М.: Медицина, 1982.- 302 с.
7. Мануилова Н.А. Гистология с основами эмбриологии. М.: Просвещение, 1973.- 214 с.
8. Троценко Б.В., Чирский Н.В. Учебное пособие по курсу гистологии (электронная версия). – Симферополь, 2001.
9. Гистология (введение в гистологию) / под ред. Э.Г. Улумбекова, Ю.А. Челещева. – М.: ГЭОТАР, 1997.
10. Гистология / Ю.И. Афанасьев, Н.А. Юрина, Б.В. Алешин. – М.: Медицина, 1989.
11. Гистология/ Ю.И. Афанасьев, Н.А. Юрина, Е.Ф. Котовский и др. – М.: Медицина, 2002. –
12. Луцик О.Д., Иванова А.И. Кабак К.С. Гистология людини. – Лвів. Мир, 1992.
ДОПОЛНИТЕЛЬНАЯ ЛИТЕРАТУРА
1. Елисеев В.Г. Гистология. М.: Медицина, 1983.
2. Куприянов В.В. и др. Сосудистый эндотелий.- К.: Здоровье. – 248 с.
3. Абрамов М.Г. Гематологический атлас. –М.: Медицина. 1985.- 237 с.
4. Шаде Дж., Форд Д. Основы неврологии. М.: Мир, 1976.- 350 с.
5. Гистология (Введение в патологию). Под ред. Э.Г. Улумбекова, Ю.А. Челышева.-М.: ГЭОГАР. 1997.- 947 с.
6. Прохончуков А.А. и др. Гомеостаз костной ткани в норме и при экстремальном воздействии.- М.: Наука, 1984.- 200 с.
7. Соколов В.Е. и др. Адаптивные свойства эпителия и его производные. Атлас микрофотографий.- М.: Наука, 1979.- 110 с.
8. Гурфинкель В.С., Левик Ю.С. Скелетная мышца и функция. –М.: Наука,1985.- 143 с.
9. Кауфман О.Я. Гипертрофия и регенерация гладких мышц. М.:Наука. 1979.- 183 с.
10. Клишов А.В. Гистогенез и регенерация тканей.- Л.: Медицина, 1984.–30 с.
11. Комиссарчик Я.Ю., Миронов А.А. Электронная микроскопия клеток и тканей.- Л.: Наука, 1990.- 140 с.
12. Заварзин А.А. Основы частной цитологии и сравнительной гистологии многоклеточных животных. Л.:Наука, 1976.
13. Гацко Г.Г. Жировая ткань при старении. Минск. Наука и техника, 1985.- 184 с.
14. Гаврилов О.К. Клетки костного мозга и периферической крови –М.: Медицина, 1985.- 288 с.
15. Вельш У., Шторх Ф. Введение в цитологию и гистологию животных. М.: Мир.1976.
16. Виноградов В.В., Воробьёва Н.Ф. Тучные клетки, Новосибирск, Наука, 1973.
17. Занябуш Л. Молекулярная и клеточная биология. М.: Мир. 1982. Т.3.
18. Зенгебуш П. Молекулярная биология. М.: Мир, т.1-3. 1982.
19. Саркисов Д.С. Очерки по структурным основам гемостаза. М.: Медицина, 1977.
20. Сборник «Молекулы и клетки». Под ред. Г.М.Франка. Выпуски 1-5. М.: Мир, 1966-1970.
21. Серов В.В., Шехтер А.Б. Соединительная ткань. М.: Медицина. 1981.
22. Касавина Б.А., Торбенко В.П. Жизнь костной ткани. М.: Наука. 1979.
23. Кассиль Г.Н. Внутренняя среда организма. М.: Наука. 1983.- 277 с.
24. Пигаревский З.В. Зернистые лейкоциты и их свойства. М.: Мир, 1978.
25. Питерс А., Палей С., Уэбстер С. Ультраструктура нервной системы. М.: Мир. 1972.
26. Шубникова Е.А. Лекции по гистологии. М.: МГУ, 1973.
27. Мозг (пер. с анг. Под ред. Симонова П.В.) М.: Мир. 1982.
28. Фриденштейн А.Я., Чертков И.М. Клеточные основы иммунитета. М.: Медицина. 1981.
Доклад по биофизике:
“Механорецепция”
1. Назначение
Для активного существования в окружающей среде, высшие организмы обладают большим количеством анализаторов, специализирующихся на разных типах воздействия: на свете (зрение), на звуке (слух), на вкус и запах (обаняние), на прикосновения и температуру (осязание), на гравитацию. Механорецепция входит в чувство осязания и вместе с вестибулярным аппаратам позволяет наиболее полно представить организму и сознанию положение конечностей и тела в пространстве.
2. Функции
Анализатор механорецепции, как и любой другой анализатор, включает три пространственно-функциональных части: рецептор (получение и трансформация механического воздействия в электрический импульс), проводниковые пути (передача импульса) и нервный центр (анализ полученной информации и формирование эфферентного ответа). Все эти части обеспечивают:
Восприятие механических воздействий на кожу: локализация, направление движения, скорость деформирующего источника, его вибрации (тактильная рецепция);
- Восприятие механических сдвигов внутри органов и мышц с целью определения положения конечностей и тела в пространстве (проприоцепция);
- Восприятие воздействий на волосяной покров.
- Трансформацию сигнала из механического в электрический, который может быть передать по нейронам.
- Передачу импульсов с высокой или низкой скоростью к нервному центру.
- Формирование в сознании общей картины положения тела и конечностей в пространстве.
- Обеспечение вегетативной нервной системы информации о положении тела и его контроль (поддержание вертикального положения тела при возможном засыпании в таком положении, информация о глазодвигательных движениях во сне).
3. Устройство и принцип работы.
Типичные механорецепторы, как правило, представляют собой инкапсулированные образования. Некоторые из них называют поверхностными концевыми органами, т. к. они в коже расположены поверхностно. Это диски Меркеля, тельца Мейснера, тельца Фатера-Пачини, тельца Догеля, колбы Краузе, тельца Руффини, нервно-сухожильные веретена, нервно-мышечные веретена и прочие (Рис.1).
Рис.1. Различные виды механорецепторов
Пластинчатые тельца (по старой терминологии тельца Фатера-Пачини)
располагаются в соединительной ткани внутренних органов и глубоких
слоях кожи, особенно на подушечках пальцев, на брыжейке, в молочной
железе, кишечнике и других внутренних органах. Имеют вид округлых
образований.
Структурными компонентами являются:
Внутренняя колба (луковица), образованная видоизмененными леммоцитами, в которую проникают нервные волокна;
- наружная колба – слоистая соединительнотканная капсула из фибропластов и коллагеновых волокон, образующих концентрические пластины, между которыми имеется жидкость.
Внутренняя часть капсулы содержит плоские, концентрически расположенные нейроглиальные клетки, ограничивающие внутреннюю колбу. Рецепторное волокно входит во внутреннюю колбу с одного его полюса и образует контакты с глиальными клетками. Терминальная часть рецепторного волокна содержит мелкие сферические митохондрии и светлые синаптические пузырьки. Внешний слой капсулы состоит из мощной соединительнотканной оболочки, образованной из плоских серповидных клеток и соединительнотканных волокон, между которыми находится интерстициальная жидкость. Пластинчатые тельца воспринимают ощущение вибрации, натяжения, давления на органы и внутриорганное давление. Принцип работы телец Фатера-Пачини на сегодня плохо изучен. Соединительнотканные пластинки и интерстициальная жидкость капсулы вероятно способствуют усилению давления на нервное окончание, в результате чего аксолемма деформируется, изменяется её проницаемость и происходит генерация потенциала. Полагают, что местом возникновения потенциала действия в тельцах Пачини является область первого перехвата Ранвье.
Осязательные тельца (Мейснера) размещаются в сосочковом слое дермы, имеют эллипсоидную форму и небольшие размеры. Это тактильные рецепторы, реагирующие на прикосновение. Присутствует в дерме кожи, особенно часто в кончиках пальцев, подошвах, сосках, веках, губах и половых органах. В центре тельца Мейснера располагается спирально свёрнутое безмиелиновое разветвление миелинового волокна, которое проходит через поперечно расположенные овальные клетки, напоминающие шванновские клетки. Снаружи тельце покрыто соединительнотканной капсулой. Внутренняя колба состоит из глиальных клеток, лежащих перпендикулярно длинной оси тельца, между которыми располагаются веточки дендритов. Снаружи находится очень тонкая, переходящая в периневрит слоистая капсула – наружная колба. Незначительная деформация капсулы передается глиоцитам и далее на дендриты.
Клетки Меркеля расположены под эпидермисом, обладают крупными неправильной формы ядрами и микроворсинками, простирающимися к эпидермальным клеткам. У их оснований располагаются дисковидные окончания сенсорных аксонов (диски Меркеля). Группа из 10 - 20 клеток Меркеля образует синаптические контакты с окончаниями одного сенсорного аксона (тельца Пинкуса-Игго). Клетки Меркеля реагируют на внезапные смещения кожи, например при поглаживании.
Тельца Руффини лежат в соединительнотканной части кожи и капсулах суставов: они воспринимают давление и имеют вид веретеновидных структур. Внутреннюю колбу образуют глиальные клетки между которыми располагаются терминали дендритов с расширениями на концах. Капсула хорошо выражена.
Колбы Краузе – мелкие округлые тельца, являющиеся механорецепторами и холодовыми рецепторами. Они лежат в дерме кожи, слизистой оболочке полости рта, подгортанника, в конъюнктиве глаза. Внутренняя колба образована плоскими глиоцитами между которыми тонкие ветви дендрита образуют сплетения в виде клубочка. Наружная капсула очень тонкая.
Генитальные тельца Догеля – находятся в особо чувствительных областях кожи (наружные половые органы, молочные железы). Они по строению похожи на колбы Краузе, но в отличие от них в тельце входит несколько отростков от нейроцитов, что вызывает сильную иррадиацию возбуждения. Реагируют на давление импульсами, вызывающими половое возбуждение.
Механочувствительные свободные окончания в коже
. Являются
немиелинизированными аксонами, у них нет корпускулярных структур.
К ним относятся рецепторы волосяных фолликул, реагирующих на давление
от движения волоса.
Нервно-мышечные веретена
– рецепторы растяжения поперечнополосатых
мышц - нервные окончания, обладающие как чувствительной, так и двигательной
иннерваций. Чувствительные нервные волокна витками обильно оплетают
несколько мышечных волокон, образуя вокруг них подобие муфты. В
этой области мышечные волокна истончаются, число миофибрилл в них
уменьшается, а количество ядер резко возрастает. Нервно-мышечные
веретёна бывают окружены соединительнотканной капсулой.
Двигательные нервные волокна образуют мелкие нервномышечные синапсы по краям интрафузальных волокон, обеспечивая их тонус, регулируя длину волокон. Всё свободное пространство между мышечными волокнами заполнено жидкостью и ограничено тонкой капсулой. Изменение тонуса мышцы ведет к изменению давления жидкости и передается на дендриты. Кольцеспиральные окончания реагируют на изменение длины мышечного волокна и на скорость этого изменения, гроздевидные окончания – только на изменение длины. Число веретен в мышце зависит от её функции и тем выше, чем более точными движениями она обладает.
Нервно-сухожильные веретена – рецепторы растяжения, располагаются в месте соединения мышцы с сухожилием, веретеновидные структуры длиной 0,5-1 мм. Каждое веретено имеет капсулу из фиброцитов, которая охватывает группу сухожильных пучков, оплетенных окончаниями нервных волокон. Возбуждение рецепторов возникает при растяжении сухожилия во время мышечного сокращения.
От рецептора импульс движется по рефлекторной дуге через спинной мозг и ствол мозга к ядрам таламуса и далее к коре.
4. Рабочие характеристики
Количество и плотность размещения рецепторов:
Общее количество тактильных рецепторов оценивается в 10 миллионов штук, сгруппированных в 1 миллион афферентов.
- Количество свободных афферентных окончаний в коже: 50% от общего количества кожных афферентов.
- Плотность размещения свободных нервных окончаний в коже: 170 шт/см2
Размеры и прочие структурные параметры:
Тельца Мейснера: 50-140 мкм
- Колбы Крауза: 40-150 мкм
- Тельца Руффини: 1-2 мм
- Тельца Фатера-Пачини: 0.5-5 мм
- Нервно-сухожильные веретёна:
- Размеры: 0.5-1 мм
- Количество мышечных волокон в веретене: 15 штук
- Нервно-мышечные веретёна
- Количество мышечных волокон в веретене: 1-8 штук
- Диаметр выходящих нервных волокон: 17 мкм (первичные) и 8 мкм (вторичные)
Скорость передачи:
Психофизическая оценка ёмкости тактильных каналов: 5 бит/c
- Скорость передачи импульсов из свободных нервных окончаний: 1 м/c (С-волокна, IV тип)
- Скорость передачи импульсов из инкапсулированных рецепторов: 50 м/c (А-волокна, II тип)
Разрешающая способность и пороги ощущений:
Порог ощущения кожных рецепторов при надавливании: 10 мкм
- Одновременный пространственный порог кожных рецепторов:
- На губах и кончиках пальцев: 1-3 мм
- На спине, плечах, бёдрах: 50-100 мм
- Последовательный пространственный порог кожных рецепторов:
- На губах и кончиках пальцев: 1 мм
- На спине, плечах, бёдрах: 10-20 мм
- Порог ощущения вибрации: 150-300 Гц
- Минимальная ощущаемая амплитуда вибрации: 1 мкм
Модальность (специфичность к воздействиям):
Тельца Руффини и колбы Крауза помимо механических воздействий детектируют температуру, т.е. полимодальны. А тельца Мейснера могут ощущать вибрацию. Также полимодальны неинкапсулированные свободные афференты.
- Остальные рецепторы унимодальны.
Рецепторные поля (площадь рецепции):
Для тактильных дисков площадь равна анатомическим размерам.
- Для дисков Меркеля это агломерация из 30-50 дисков, обслуживаемых одним нервным волокном.
- Для телец Фатера-Паччини эта область больше анатомически размером.
Адаптация (см. Таблица 1).
Таблица 1. Классификация кожных механорецепторов по скорости адаптации и адекватным стимулам.

5. Регуляция
Информация от рецепторов передается в центральную нервную систему
через спинальные или черепномозговые нервы. Ответвления аксонов,
передающие сенсорную информацию, формируют синаптические окончания
на нейронах спинного мозга (или ствола мозга в случае черепномозговых
червов), затем направляются в высшие отделы нервной системы (стволовые
ядра, которые в свою очередь передают информацию выше). В каждом
из этих отделов нервной системы поток сенсорной информации может
фильтроваться - акцентрироваться или, наоборот, блокироваться.
Сенсорные системы построены по иерархическому принципу: сигналы от рецепторов поступают в низшие уровни центральной нервной системы (спинной мозг или ствол мозга), откуда передаются в более высшие отделы (ядра таламуса, кора больших полушарий, базальные ганглии). На каждой из этих последовательных стадий сенсорная информация преобразуется, происходит ее фильтрация.
Поток информации не односторонний, так как высшие отделы иерархии
посылают сигналы в низшие отделы. Кроме того, сенсорная информация
обрабатывается не цепочкой последовательных структур, а, скорее,
многими областями мозга одновременно (или, как говорят, параллельно).
Параллельная обработка сенсорной информации становится очевидна
в высших областях мозга, таких как кора полушарий. Здесь отдельные
области специализируются на обработке отдельных элементов информации.
Сенсорная информация различного рода не обрабатывается отдельно.
Во многих областях мозга, называемых ассоциативными, происходит
смешение модальностей - например, в париетальной коре и буграх четверохолмия.
Нейроны в этих отделах мозга реагируют на различные стимулы, например,
зрительные, тактильные и слуховые.
Структурами, контролирующими сенсорную информацию, являются кора
больших полушарий (в частности, префронтальная кора, играющая важную
роль в контроле внимания), базальные ганглии, ретикулярная формация,
таламус (в частности, ретикулярное ядро таламуса) и другие структуры.
Одним из основных механизмом фильтрации сенсорной информации является торможение, которое производят ГАМК-эргические синапсы. Как правило, нейрон передающий сенсорную информацию, является возбуждающим. Фильтруют эти возбуждающие сигналы тормозные нейроны. Торможение может быть пресинаптическим (то есть блокирующим передачу сигналов по сенсорному аксону к какому-либо нейрону) либо постсинаптическим (гиперполяризующий нейрон, принимаюший сенсорные сигналы). Постсинаптическое торможение позволяет блокировать сигналы избирательно, так как принимающая клетка остается способной отвечать на другие, незаблокированные воздействия.
На фильтрацию сенсорных сигналов также оказывают влияние такие нейромедиаторы как ацетилхолин, дофамин, эндорфины и другие.
6. Энергозависимость
Так как весь анализатор механорецепции является производным нервной ткани, то энергозависимость особенно высокая. Нейроны снабжаются питательными веществами и кислородом через нейроглию и расположенные в ней кровеносные сосуды. При гипоксии и тромбах может нарушаться питание отдельных участков (например кожи или органов), начнётся ретроградная дегенерация нейронов (в некоторых случаях возможно восстановление функций волокна через некоторое время после восстановления питания) и утрату рецепции на участке. Однако гипоксия в мозге может привести к гораздо худшим последствиям, утратой функций целых отделов или полей коры мозга.
7. Технический аналог
Аналогом механорецепторов являются устройства, основанные на пьезоэффекте. Тельца Руффини в своём составе имеют коллагеновые волокна, а они проявляют пьезоэлектрические свойства. Пьезоэлектрический эффект (пьезоэффект) состоит в том, что при механических деформации некоторых кристаллов в определённых направлениях на их гранях появляются электрические заряды противоположных знаков, т.е. механические воздействия трансформируются в напряжение. Получается, что тельца Руффини функционируют как своеобразные пьезоэлектрические устройства, поскольку в них между нервной терминалью и коллагеновым волокном нет какой-либо иной ткани. В технике часто используется обратный пьезоэффект, когда электрический ток преобразуется в механические колебания – например, для генерации ультразвука.
Список литературы
1. Шубникова Е.А., Функциональная морфология тканей. – М: Издательство
Московского университета. – 1981. – 326 с.
2. Фенькина Р.П., Дегтярев В.П., Коротич В.А., Учебное пособие по
нормальной физиологии. – М:МГМСУ. – 1994г
3. Сандаков Д.Б., Курс лекций по физиологии. – Минск: биофак БГУ.
4.
Шмидт Р., Основы сенсорной физиологии. – М:Мир. – 184. – 287 с.
5. Кузнецов С.Л., Мушкамбаров Н.Н., Горячкина В.Л. Руководство-атлас
по гистологии, цитологии и эмбриологии.

Если вы задумались над вопросом о том, чем бы новым и интересным занять вашего маленького мастера, то предлагаем вам...
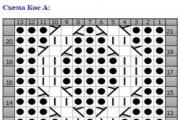
выглядят очень нарядно и модно. Но выбирая интересный фасон для ребенка, обязательно надо учитывать качество пряжи , из...

Привет, ребята! Продолжаю тему 8 марта, а именно хочу поделится своими находками красивых и в тоже время не слишком...

Самодельные игрушки пользовались популярностью всегда. Нельзя считать этот вид игрушек устаревшим, ведь он приносит много...

Без любви невозможно полноценное счастье. К сожалению, не у всех получается найти любовь и сохранить ее на долгие годы....

Эфирное масло чайного дерева - светло-жёлтая или бесцветная жидкость, которой присущ весьма специфический, слегка...
Женщины, планирующие родить ребенка, сталкиваются со множеством незнакомых медицинских терминов. Их значение...

Полезные советы Наступило время, когда все мы начинаем утепляться и вытаскивать свитеры из шкафа. Но если вы...

Пико, связанное крючком (или, как его еще называют, пике) – это самый простой, но и в тоже время, красивый узор...

Geox – бренд, подаривший миру дышащую обувь Итальянская компания Geox создаёт и производит верхнюю одежду и...

Одна из самых красивейших и любимых сказок детства «Снежная королева». Многие из нас по нескольку раз...

Содержание Введение………………………………………………………… ………………3 Глава 1. Теоретические основы проблемы организации досуга...

Мастерицы предпочитают начинать свое обучение вязанию крючком с воплощения простых схем салфеток. К сожалению,...

Весна - самое время пополнить свой гардероб яркими и красивыми новинками. Талантливый модельер поделился...
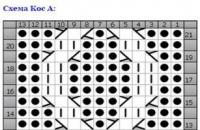
выглядят очень нарядно и модно. Но выбирая интересный фасон для ребенка, обязательно надо учитывать качество...

Привет, ребята! Продолжаю тему 8 марта, а именно хочу поделится своими находками красивых и в тоже время не...